Atmospheric image shot on camera trap of AI (Image Keith Armstrong)
Building on this prior post that sought to accelerate the decomposition of the Art Intelligence/hybrid installation on site – its salient to remember that each AI must somehow ‘evolve with’ and ‘learn from’ the emerging forest whilst directly enhancing forest’ fluxes of intelligent natural regrowth.
How to connect the intelligence + accelerator as a proxy for the rest of the forest (.e. Ads are placed at principal, representative sites, to stand in as proxy/exemplar for the entire forest development)
Undertake mapping survey of nascent trees in the vicinity – recalling that the log was placed within a patch of non-native noxious Paspalum notatum (Bahia) grass – with the intention it might assist its decline back to native vegetation.
LiDAR scan the log (to add to the recently scanned model of that site (under construction))
Construct a visual representation of the relationships (refer here to literal/chemical/speculative/imporobable links – referring to this recent book, Light Eaters)
AI Acceleration Elements
Define sections of the tree where different eco-spatial, acceleration experiments will take place, mapping out in advance:
Construct (laser cut) flexible template for drilling the tree to encourage decomposition: may include point cloud LiDAR model holes – mapped to tree’s physical decay holes (potentially use high torque of portable angle grinder to increase cut efficiency).
Add other fallen debris and forest floor organism to the vicinity (Some from original fallen limb site and some from adjacent forest to begin to develop enhanced ‘bridges’ between growth)AI Interpretation Elements
Establish possible components of the Art Intelligence Interpreter outcomes from these. (Art Intelligence interpreters (Aii) = Elements that create additional layers of engagement with the Ais, and their hosting forest, intended predominantly for human audiences (art and otherwise).
Establish outdoor cameras around tree to monitor
– Drilled positions – capable of monitoring fugal evolution
across decay hole structure night and day
– relative impacts on nearby vegetation (wrt to the whole plot).
Install reliable phenocams across the whole plot
Add degradable rolled up visual images to each hole and monitor their relative decay
Consider adding tiny solar lights within some holes to encourage local nighttime insect activity
Install the Audiomoth sound monitor
Install camera trap to further understand wildlife movements
Art Intelligence accelerators (Aia) = Additional/embellishing, creative elements added to Art Intelligences to enhance and accelerate local ecological processes – therefore intended primarily for non-humans. For example, these may add additional benefit or encouragement to certain organisms to be and become, that in turn will further aid forest recovery.
Art Intelligence on site, Nov. 2024 (Image Keith Armstrong)
The benefits to the forest of introducing coarse woody debris Coarse woody debris (like the two-tonne log introduced to the site) is critical to functioning of an ecosystem. In a paddock or on a woodland floor, such grounded timber stabilises soil, reduces erosion, and creates a protected microclimate where small animals can thrive, nutrients collect, and seedlings can gain a foothold protected from grazing and the elements. Hundreds of species of wildlife rely on fallen timber for shelter and food resources, (while standing dead trees often provide hollows that are important nesting sites for wildlife). It can take decades, to centuries to accumulate – especially larger pieces. It has been lost from many ecosystems (such as the artwork site at SERF) globally due to veg clearing, logging, forest regimen and grazing. At the artwork site SERF (Qld) the paddock had been bare for decades, and only an introduction as we have just done would bring back such debris.
FAI – Fallen Tree, Nov 2024, (Image Keith Armstrong)
Ecology is a charmed science. It provides us with the capability, the acuity, to see beyond the simple veneer of a place or environment and instead perceive another dimension: the convoluted interplay of its components, the depth of its connections, its patterns and processes. To do ecology is exhilarating; it is transformative; it is about becoming something else, about shedding the skin and perspectives that clutter and distort our human-centric myopia; to do ecology is about pricking the delusion of being apart from and above nature. It is the wonder of seeing the world from the perspective of different species. It is about understanding the endless and enduring shifts of time and place, the infinitely variable patterning inherent in the natural world, in the world into which we must fit. Woinarski, J Z (2023) To the future: An ecology of love, hope, and action. Austral Ecology 48: 1705–1712.
Retaining fallen timber in different environments throughout the landscape supports more complex and resilient ecosystems. Removing fallen timber interrupts all of these processes, leading to reduced biodiversity and less resilient ecosystems. It can also have a devastating effect on wildlife. The benefits of coarse woody debris include:
• developing soil by cycling nutrients back into the soil as the timber degrades
• Reducing erosion and stabilising slopes and gullies
• Supporting more productive microclimates and helping retain localised soil moisture through water infiltration and improved thermal conditions.
• Creating a site for re-colonisation by ground cover plants, particularly wind and water dispersed plants, following fire or other disturbance
• Providing shelter, habitat, food resources and foraging grounds for native species, from tiny microbial communities through to birds and mammals. (Fallen timber and dead trees can act as stepping
stones for fauna and provide a substrate and nutrients for invertebrates and fungi). https://www.sustainablefarms.org.au/wp-content/uploads/2021/01/Fallen-timber-and-dead-trees-ONLINE_0.pdf
FAI – Fallen Trees, Nov 2024 (Image Keith Armstrong)
But to do ecology – and especially conservation biology – in this age is also devastating. Our science, our understanding, uniquely sharpen our perspectives, give us a cursed privilege of seeing the cracks, about watching them widen, about feeling grief at the casualties, about knowing that the future will likely be characterized by increasing loss. Woinarski, J Z (2023) To the future: An ecology of love, hope, and action. Austral Ecology 48: 1705–1712.
Our partners TERN use protocols for measuring coarse woody debris like the log introduced to the site – ie the future site for microbes of all kinds which project collaborator Dr. Eleanor Velasquez drew my attention towards (and before that Dr. Lebbinck too in this post here).
Typical approaches to restoration involve plant only and plant and animal only approaches – which operate under what the authors call the ‘field of dreams paradigm’ – i.e. that if you build the habitat they will come! Such attempts often therefore ignore the ‘unseen majority’. Contos P, Wood JL, Murphy NP, Gibb H. Rewilding with invertebrates and microbes to restore ecosystems: Present trends and future directions. Ecol Evol. 2021;11:7187–7200. https://doi.org/10.1002/ece3.7597.
Here is the way TERN classify the stage of rot of materials.
Coarse woody debris decomposition/ Art Intelligence accelerators (Aia)
The next step is to determine potential Art Intelligence Accelerator processes that will expedite the decay of the log from what TERN would call Class 1 (Recently fallen/structurally intact) to a higher class (e.g. cannot support its own weight/soft to kick).
Fallen tree branches such as ours don’t rot quickly as they’re almost all carbon. To compost properly, the requisite microbes/fungi need nitrogen, and other trace nutrients. (The ideal ratio of carbon to nitrogen in compost is around 30 to 1. That’s 30 parts carbon for every 1-part nitrogen. Wood is naturally around 500 to 1, since it’s almost entirely carbon). Microbes therefore need a lot more nitrogen to break down a stump and so acceleration of the decomposition processes will assist building the soil and health of the re-forestation site.
One approach involves increasing the surface area so it’s more exposed to the elements (e.g. by drilling or slotting the log), while at the same time feeding the bacteria and fungi that will decompose it. Mulching the log also helps hold in moisture and encourages the stump to break down faster.
Art Intelligence/Coarse Woody Debris on site, Nov. 2024 (Image Keith Armstrong)
Key techniques include:
Drilling holes (or chainsaw slots etc) into the wood helps it retain water – via little pockets that trap rain, moisture, and nutrients – and attract insects who will then chew additional holes, breaking it into smaller pieces that decay faster. The water caught in those pockets also promotes fungus growth – and given that water helps the wood rot more effectively, making sure the stump stays consistently moist is important.
Sprinkling fungus spores or pieces of grown fungus onto the wood collected from fallen logs near the area, ideally from rotting wood of the same type, is also a viable approach. (Many fungi species are generalists and so any spores applied will likely help).
Spore transfer (Image https://www.hunker.com/13407080/how-to-speed-up-wood-decay/)
Another key approach is covering the wood with sod or peat moss or nearby soil from the existing forest (thereby inoculating the site with microbes) -to keep it moist and its temperature stable. A moisture content of 20 percent is optimal and temperatures of 10 to 32 degrees C promote fungus growth. NB sunlight is not required because fungi do not photosynthesise. In our case this will mean fungal growth is more likely in the cooler months given the forest has yet to really take off.
Back to soil (Image https://www.hunker.com/13407080/how-to-speed-up-wood-decay/)
Adding nutrients is also valuable – i.e. applying a one off small amount of high-nitrogen fertilizer to the ground near the edge of the stump or log to kick-start the fungus growth – e.g. Miracle Grow (or other high nitrogen fertilizer) works well, but is not as concentrated as potassium nitrate. Pure potassium nitrite pellets are pelletized nitrogen fertilizers. Lab-grade potassium nitrate is more natural than a such prepared blends that includes other potential toxins. “Yeast nutrient,” is a food-grade nitrogen that’s used in winemaking (not the same as nutritional yeast, and it can only be purchased online or at specialty winemaking or homebrewing stores) or use other things you have on hand, like natural cultured buttermilk (contains probiotics that will help digest the stump but much slower than other nitrogen sources), or urine – by peeing on it regularly. NB the least effective method, is urine doesn’t have nearly as much nitrogen as actual fertilizer.
Epsom salt will also encourage rot to some extent – as it is a magnesium salt, and many microbes do require magnesium, at least in low levels. However it’s not the main limiting nutrient though, and it won’t be nearly as effective as adding Nitrogen.
Pouring granulated sugar into the holes and nooks of the wood after one year has passed since the fertilizer application. The sugar provides an additional carbon source for the fungi. Repeat this step every few months until the wood has completely decayed.
Myrtaceae(/mərˈteɪsiˌaɪ, -siːˌiː/) Science/The Site
“As they slowly come together as a renewed community, they will be an extraordinary and invaluable addition to our Australian heritage.”
Dr. David Tucker (10/8/24, EF grant reference letter)
SERF Myrtacea, Sept 2024 )(Image Keith Armstrong)
“Clearly Eucalypt species such as these are in effect ‘hiding in plain sight’ in the Australian public’s imagination, and I have long asked how we might encourage people to begin to better see what we risk losing. Clearly, we need other approaches beyond science to overcome our ‘plant blindness’, and this is why I support the Forest Art Intelligence project’s aim of bringing attention to the described natural intelligence of this Eucalypt woodland as it recovers over time”.
Dr. David Tucker (10/8/24, EF grant reference letter)
The purpose of this post is to attempt come to come to terms, maybe just a little more, with some of the artwork sites’ eucalypt species given the project is focused upon their flourishing. This will both enhance the shared language with the science team, and further my capacity to differentiate the ‘wood from the trees’ whilst on site 😉 This, based upon a realisation how little I comprehend in a systematic/science0-eyed sense when in the bush, despite the all consuming experience it always engenders. Is it therefore possible to absorb more?
The predominant trees at the two artwork sites are in the order Myrtaceae {the Myrtle family} – a large, cosmopolitan family of plants with over 5000 species worldwide, well represented in Australia contain ca. 12 families with over 70 genera and over 1500 species – can be found in a wide range of habitats and climatic zones – coastal heaths, temperate forests and woodlands, tropical rainforests – even arid and alpine zones.. (https://anpsa.org.au/genera/myrtle-family-myrtaceae/).
All myrtle species are woody, contain essential oils, and have flower parts in multiples of four or five. The leaves are evergreen, alternate to mostly opposite, simple, and usually entire (i.e., without a toothed margin). The flowers have a base number of five petals, though in several genera, the petals are minute or absent. The stamens are usually very conspicuous, brightly coloured, and numerous.
The eucalypts – within the plant family Myrtaceae – number among their relatives such well known Australian genera as Callistemon (bottlebrushes), Melaleuca (paperbarks), Leptospermum (tea trees) and Syncarpia (turpentine)
Apple – A name used by early European settlers due to a similarity in appearance of some plants to apple trees (eg. Angophora bakeri, Narrow-leaved apple)
Ash – Timber is similar to the European ash trees (eg. Eucalyptus regnans, Mountain ash)
Blackbutt – The lower part of the trunk has persistent bark which is usually black due to past fires (eg. Eucalyptus pilularis, Blackbutt)
Bloodwood – Timber often has pockets of a dark red gum known as kino (eg. Corymbia eximia, Yellow bloodwood)
Box – Bark is retained on the tree and is short fibred; plates of bark may shear off with age (eg. Eucalyptus melliodora (Yellow box)
Ironbark – Bark is retained on the tree and is hard and deeply furrowed (eg. Eucalyptus crebra, Narrow-leaved ironbark)
Mallee – Multi-stemmed trees, usually fairly small in height (eg. Eucalyptus albida, White-leaved mallee)
Peppermint – The oil in the leaves has a peppermint-like aroma (eg. Eucalyptus dives, Broad-leaved peppermint)
Ribbon Gum – Bark is deciduous and is shed in long strips which often hang from the branches (eg. Eucalyptus viminalis, Ribbon gum)
Scribbly Gum – Bark is deciduous and the smooth trunk is marked with “scribbles” caused by an insect larva (eg. Eucalyptus sclerophylla, Scribbly gum)
Stringybark – Bark is retained in long fibres which can be pulled off in “strings” (eg. Eucalyptus eugenioides, Thin-leaved stringybark)
Eucalypt Bark Types
We will encourage the transition process on this plot via selective slashing, mulching, weeding, and the introduction of fallen habitat trees & occasional selective planting under Marcus Yates management and with advice and input from Dr David Tucker, Marcus Yates, Dr. Gabrielle Lebbink and Dr. Eleanor Velasquez.
SITE 1: 7164 sq. m passively managed plot – grassed sloping bank, last slashed July 23. Likely similar to the other dominant veg at SERF – i.e. Myrtaceae woodland on Mesozoic to Proterozoic igneous rocks – specifically Eucalyptus tereticornis, Corymbia intermedia, E. crebra +/- Lophostemon suaveolens woodland on Mesozoic to Proterozoic igneous rocks . This vegetation is ‘of concern’ Regional Ecosystem classification RE 12.12.12: @ 2021 – only 21.5% of this type remains as it is extensively cleared for pasture.
Re-growth on Site 1, 2024, (Image Keith Armstrong)
Emergent species: Eucalyptus crebra: Narrow-leaved red ironbark
(Eucalyptus tereticornis, (blue gum /forest red gum/red irongum),
Corymbia intermedia (pink bloodwood),
Corymbia tesselaris (Moreton Bay ash))
Reference Trees at Barracks (in process learning!)
Reference tree positions @ Barracks (Image Keith Armstrong)
Other canopy/relevant local species we had initially planned to plant Melaleuca quinquenervia (broad-leaved paperbark, paper bark tea tree),
Melaleuca salicina (willow bottlebrush),
Guoia semiglauca (guioa or wild quince)
with the expectation that many subcanopy and shrub species will passively regenerate through the dispersal of seeds from birds, water, wind etc. following canopy closure and site capture
SITE2: 7035.658 sq. m ‘wetland’, holding area – grassed seasonal wet gulley area – was burnt in August 2023 – likely an ecotone associated with wet gullies RE12.3.6, which reflects the forest type further along the drainage line – Melaleuca quinquenervia +/- Eucalyptus tereticornis, Lophostemon suaveolens, Corymbia intermedia open forest on coastal alluvial plains
Predominant weedy grass species in SERF active regeneration area gulley, Summer 2024 (Image Keith Armstrong)
To the uninitiated, most eucalypt species tend to look similar, and while taxa in some groups are indeed difficult to distinguish, in general there are good features and clear characteristics to use in identification. Eucalypt leaf morphology provides a range of diagnostic features (as well as injects a level of confusion in the change from seedling to juvenile to sapling to adult leaves that takes place in the majority of species). Eucalypt fruits (gumnuts) also show great diversity in form and size. Identification in EUCLID for eastern Australian species usually fall back on the less conspicuous and accessible but highly diagnostic characters, often ones that may be less relevant in other plant groups.
In working with eucalypts in the field it is important to recognise whether the trees are cultivated, or occur naturally. If cultivated, they could be from anywhere in Australia. To aid identification take into account other aspects of the specimen, viz. the height of the plant, the number of stems or trunks, the colour of the crown, the overall appearance of the crown to determine if it is composed of juvenile or adult leaves, general size of the leaves (very small, e.g. E. parvula or E. kruseana, or very large, e.g. E. globulus) and the type of bark, basically, whether rough or smooth, and extent of coverage by the rough bark of the smaller branchlets. There is often considerable variation in some characters between trees of the same species in one population, especially in size of parts, such as length and width of leaves, length of petioles, bud sizes, lengths of peduncles and pedicels, and fruit dimensions and position of the disc relative to the rim of the fruit.
The ‘internal’ features of the eucalypt plant, such as the number of opercula in the bud, arrangement of stamens, number of ovule rows and seed shape, are usually more reliable for identification than the ‘external’ features. They are relatively protected from the elements and from various forms of predation. They are the parts that require handling and close inspection or even dissection, as opposed to macro observation.
Calistemon in Flower, SERF, 26/9/24 (Image Keith Armstrong)
Specimens for study may be obtained.. For most trees, a weighted length of rope can be thrown over a low branch which can then be broken off with a sharp tug and pulled to the ground for close inspection of the parts. Alternatively, for trees of moderate height, pole pruners can be used less destructively than the weighted rope. For tall trees it is a curious fact that the flowers and fruits are small and scarcely visible to the unaided eye, e.g. E. regnans. Then the canopy needs to be inspected with binoculars and a useful branch selected. If it is above rope-throwing height, the branch may be reached with the use of a shanghai by shooting a lead weight attached to a fine, light line over the branch and then attaching a thicker, stronger rope to one end of this line and then pulling this line up over the branch. Often the smallest trees or mallees have the largest buds and fruits, e.g. E. pyriformis. These plants are the easiest to sample, examine and assess.
The whole process of identification begins in the field with broad external assessment and ends with microscopic examination. In summary it might be said that the number of opercula on the developing flower bud is of absolute reliability, staminal inflexion, ovule row numbers and seed shape are of high reliability, bud numbers, flower colour and bark type of medium reliability, leaf colour of low reliability, bark colour of very low reliability. External features are very susceptible to seasonal and intra-population variability.
This is the level of detail to expect to shift through scientifically:!
(Pink Bloodwood) – Myrtaceae: Tree to 35 m tall. Forming a lignotuber. (ie embedded vegetative buds that allow regeneration following crown destruction – forming new stems/trunks (which can become massive) after fire possessing
Bark rough and tessellated to the small branches, often thick and furrowed, grey or brown
Juvenile (coppice or field seedlings to 50 cm): stem rounded in cross-section, scabrid at first, becoming smooth; juvenile leaves always petiolate, peltate for ca 10 nodes, opposite for 3 to 7 nodes then sub-opposite to alternate, lanceolate, 8–16 cm long, 2–4.2 cm wide, base of lower leaves peltate, eventually tapering to petiole, discolorous, green, scabrid at first particularly on petiole and midrib, eventually smooth.Adult leaves alternate, petiole (0.8)1.2–2.5 cm long; blade lanceolate, 8–15 cm long, 1.3–4 cm wide, base tapering to petiole, discolorous, glossy or dull, green, strongly penniveined, densely to very densely reticulate, intramarginal vein parallel to and just within margin, oil glandsisland or absent. Inflorescenceterminalcompound, peduncles 1–2 cm long; buds 7 per umbel, pedicels 0.2–1.5 cm long. Mature budspyriform to obovoid, 0.7–1 cm long, 0.4–0.6 cm wide, green to yellow or creamy, smooth, without ridges, scar absent (both opercula shed together at flowering), operculumconical to rounded to beaked, stamensinflexed, antherscuboid or cuneate, versatile, dorsifixed, dehiscing by longitudinal slits (non-confluent), style long, stigma tapered or mop-like, locules 3 or 4, the placentae with ovules not arranged in distinct vertical rows. Flowers white. Fruitpedicellate (pedicels 0.2–1.5 cm long), ovoid to barrel-shaped, the neck short, 1–2 cm long, 0.8–1.6 cm wide, appearing speckled, discdescending, valves 3 or 4, enclosed. Seeds brown or reddish brown, (4)6.5–9(11) mm long, ellipsoidal with terminal wing, surface smooth, hilumclose to one edge, ventral.
Introduction to FOREST RESTORATION APPROACHES
Ecologist Susan Simmard (famous for defining the wood wide web) reminds how plants in effect suck up sunlight – photosynthesising via leaves (and sometime stems) – shuttling energy down into roots – and how they share carbon with other trees (e.g. interdependence). Tree roots and soil are the foundation of the forest – . The mycelium from the mychorizza – infects and colonises roots – and trade carbon for nutrients, forming nodes and links/fungal highways – mother or hub trees nature the young – sending carbon to their seedlings in the understory. Forests Simmard reminds us aren’t just collections of trees but complex systems with hubs (hub trees) and networks .. that overlap and allow them to communicate and provide avenues for feedback and adaptation – that make the forest resilient. In the case of FAI – that network is in its infancy/simplified and needs nurture.
Eucalypts form symbioses with mycorrhizal fungi both in their native habitat and in plantations. For instance, one study of sporocarps under Eucalyptus globulus in both plantation and forest settings reported 44 putative ectomycorrhizal (ECM) species; 30 of which occurred in plantations (Lu et al. 1999)
Inter-plant communication through mycorrhizal networks mediates complex adaptive behaviour in plant communities.
Likewise re: sentience/sapience (Sentience: The ability to feel emotions, have a subjective experience, develop a personality, and form a morality.Sapience: The ability to act rationally, to learn, to understand) – Peter Wohlleben (The Secret Life of Trees) makes compelling cases for communication and cooperation among plant species.
Please note that this info on restoration below may not all apply to a subtropical climate.. (Thanks Dr. Gabrielle Lebbinck for this heads up) Approaches to Restoration: Reestablishing native grassy swards in degraded grassy white box woodlands
( Prober et al. 2005). + (Ref 2)A critical stage in the restoration and persistence of native plant composition and diversity is seedling germination and establishment. The seedling stage is often exceptionally vulnerable and determines the distribution of many species. (Harper 1977). Consequently, conditions for seedling recruitment will influence the composition and diversity of restored plant communities (Grubb 1977; Hobbs and Huenneke 1992; Morgan 2001; Clarke and Davison 2004). Restoration of ecological conditions that promote germination and establishment of native seedlings is thus an important component of successful restoration of species composition and diversity.
SERF Forbs (Image Keith Armstrong)
FORBS
Temperate grassy eucalypt woodlands in the agricultural zones of south-eastern Australia naturally support a high diversity of herbaceous perennial forbs among the dominant grass tussocks (McIntyre et al. 1993; Prober and Thiele 1995; Clarke 2000). These ecosystems have become highly fragmented and degraded through nearly 200 years of clearing and agricultural use, and most woodland remnants now have a high abundance of exotic plants and reduced native plant diversity (Lunt 1991; Tremont and McIntyre 1994; Prober and Thiele 1995). Ecological restoration of understorey plant diversity is thus urgently needed to conserve and enhance what remains of these grassy ecosystems (Cole and Lunt 2005; Prober and Thiele 2005; Gibson-Roy et al. 2007), and to ensure the long- term survival of many woodland forb species.
However, restoration of native plant diversity in temperate grassy ecosystems is not straight forward(McDougall and Morgan 2005). Understorey degradation in these ecosystems has been associated with altered ecological processes, particularly enhanced soil nutrient regimes (McIntyre and Lavorel 1994; Prober et al. 2002a; Dorrough et al. 2006). These can favour the long-term persistence of exotics (Prober et al. 2002b, 2005) and modify conditions for establishment and survival of many native species. In particular, competition from exotics may reduce resources for seedling establishment by native species (Grime 1979; Wilson and Tilman 1993; Lenz and Facelli 2005) and suppress slower-growing native species (Alvarez and Cushman 2002; King and Buckney 2002; Prober et al. 2002b).
Oversupply of red natal weedy grass, SERF plot, Sept 2024 (Image Keith Armstrong)
Restoration techniques that have attempted to address these altered ecological processes include spring burning, seasonal grazing and carbon addition. Spring burning removes litter, above-ground biomass and the seed bank of exotic annual grasses (Kost and De Steven 2000; Clarke and Davison 2001; Prober et al. 2004, 2005), reducing the abundance of annual exotic grasses in the following seasons, but variously enhancing broadleaf exotics (Prober et al. 2005). Heavy grazing in spring has similarly been found to reduce the seed set and abundance of exotic annual grasses (Menke 1992; Garden et al. 2000). Carbon addition promotes carbon-limited soil microorganisms, which subsequently compete with plants for available soil nitrogen, dramatically inhibiting the growth of nitrophilic annual exotics and in some cases allowing slower-growing native species a window-of-opportunity to establish (Averett et al. 2004; Corbin and D’Antonio 2004; Prober et al. 2005). Spring burning and carbon addition have proven successful for establishing native grasses in Australia (Prober et al. 2005) and grasses and forbs in tall-grass prairies in the USA (Baer et al. 2003; Blumenthal et al. 2003; Averett et al. 2004), and have led to successful restoration of ecological processes associated with nitrogen cycling in temperate grassy eucalypt woodlands (Prober et al. 2005).
An important next step in restoring woodland understoreys is to enhance native plant diversity by re-establishing sustainable populations of native perennial forbs. However, there is little data on natural recruitment and population dynamics of native forbs to guide this process (Morgan 2001; Clarke and Davison 2004), and responses of native forbs to the above restoration techniques (especially carbon addition) are unknown.
———————- TAXONOMY ADDENDUM —————–
https://en.wikipedia.org/wiki/Taxonomy_(bi
LIFE: Eukaryota – Organisms whose cells have a membrane-bound nucleus.
DOMAIN/CLADE: Diaphoretickes – The majority of the earth’s biomass that carries out photosynthesis belongs to Diaphoretickes – 400,000 members.
KINGDOM/PLANTAE – predominantly photosysnthesisers who obtain their energy from sunlight, using chloroplasts derived from endosymbiosis with cyanobacteria to produce sugars from carbon dioxide and water, using the green pigment chlorophyll. (Exceptions are parasitic plants that have lost the genes for chlorophyll and photosynthesis, and obtain their energy from other plants or fungi. Most plants are muliticellular, except for some green algae. NB Plantae excludes fungi and some algae.)
PHYLUM/Traceophytes -Vascular plants (from Latin vasculum ‘duct’), also called tracheophytes (UK: /ˈtrækiːəˌfaɪts/,[5]US: /ˈtreɪkiːəˌfaɪts/)[6] or collectively tracheophyta. are plants that have lignified tissues (the xylem) for conducting water and minerals throughout the plant. They also have a specialized non-lignified tissue (the phloem) to conduct products of photosynthesis. The group includes most land plants (c. 300,000 accepted known species)[10] other than mosses
CLASS/Angiosperms – Flowering plants are plants that bear flowers and fruits, They include all forbs (flowering plants without a woody stem), grasses and grass-like plants, a vast majority of broad-leaved trees, shrubs and vines, and most aquatic plants.
ORDER/Eudicots – flowering plants (angiosperms) which are mainly characterized by having two seed leaves (cotyledons) upon germination.[1]The term derives from dicotyledon (etymologically, eu = true; di = two; cotyledon = seed leaf).
FAMILY/Rosids – Today’s broadleaved forests are dominated by rosid species, which in turn help with diversification in many other living lineages. Additionally, rosid herbs and shrubs are a significant part of arctic/alpine and temperate floras. The clade also includes some aquatic, desert and parasitic plants
Malvids
The malvids consist of eight orders of flowering plants: Brassicales, Crossosomatales, Geraniales, Huerteales, Malvales, Myrtales, Picramniales and Sapindales.
The Myrtaceae genera – https://www.researchgate.net/figure/Plastome-phylogeny-of-Myrtales-based-on-an-ML-analysis-of-78-genes-from-the-plastome-of_fig2_342821472
On June 11th 2024, almost a year after the first (winter 2023) plant survey, Dr. David Tucker and Dr. Gab Lebbink undertook the second botanical survey on site – using three established transects that crossed the passive regeneration artwork site, and one end of the newly named wetland artwork site. The vegetation types had now shifted from a recently mowed winter site back in 2023 (following a dry season and pre-burn) to a now actively regenerating area after extensive rain – which had also long recovered from its winter 2023 burning.
We surveyed the site along the three prior transects – with each of the 5 sites along each transect marked by a ground peg installed in 2023.
Plant transects and quadrat sites for botanical survey (approx.) (Image Keith Armstrong)Botanical Survey transect (with tape measure on the ground) running across survey site, June 11th, 2024 (Image Keith Armstrong)
An ecologists’ quadrat (1x 1m) was laid at each pegged point (see diagram above) and estimates of ground cover (% ) and lists of species recorded. As before the agreement on the floristic content was negotiated through a wonderful, gentle dialogue between the two ecologists – a soundtrack I again recorded a part of for use in a future artwork – just as I had used it the artwork Analog Intelligence. Next a 5x5m square, originating from that quadrat point at each peg was surveyed for tree growth – with species names, counts and heights recorded. These figures therefore gave us an estimate across the entire site of vegetation type, quality and mix.
Typical bare plot from initial survey in June 2023 – with young trees marked with tape (Image Gabrielle Lebbink)Typical rich plot from second survey in June 2024 – Dr. Gabrielle Lebbink laying down a quadrat at the second site plant survey, June 11th, 2024 (Image Keith Armstrong)A plethora of life in the renewing forest/grassland (Image Keith Armstrong)
As a result, the plots were each quite dense and broadly speaking the number of trees had generally grown well (although some had been lost in the 2023 burn, the very wet summer – or possibly were also hidden in the long grasses and forbs).
Re-growth of trees now evident everywhere at the site, Botanical survey day, 11/6/24 (Image Keith Armstrong)Dr. David Tucker and Dr. Gabrielle Lebbink working in the field at the second site plant survey, June 11th, 2024Dr. Gabrielle Lebbink at the second site plant survey, June 11th, 2024 (Image Keith Armstrong)A plethora of life in the renewing forest/grassland, June 2024 (Image Keith Armstrong)
In due course, the results of the survey will be collated and compared – which will tell us about the practical progression of the project’s science/ecological meta-aims – i.e. to regrow a native forest from a cleared block – whilst allowing the natural intelligence of the forest and all its constituent species to direct that progress, and with minimal intervention.
Scene from the artwork ‘Analog Intelligence, 2024’ that suggests the nascent forest returning to the artwork site (Image Keith Armstrong)
Comparative Surveys
Here is how the site had progressed ny 11/6/24 (or regressed in some cases since the initial survey – which was followed by a burn). There had certainly been some significant changes with some problematic weeds decreased ) e.g. Paspalum urvillei – and at that stage the field still had end of the epic native Fimbristylis spp. crop!
Life Form Key (Perrenial/Annual P/A Graminoid/Forb G/F. Shrub/Climber S/C)
#
Species
Provenance
Life
Form
1707
23
1106
24
1
Cynodon dactylon
Exotic
PG
Y
Y
2
Digitaria spp.
Exotic
PG
Y (*1)
Y
3
Centella asiatica
Native
PF
Y
Y
4
Hypochaeris radiata
Exotic
PF
Y
Y
5
Imperata cylindrica
Native
PG
Y
Y
6
Fimbristylis spp.
PG
Y
N
7
Ageratum houstonianum
Exotic
PF
Y
Y
8
Sporobolus spp creber
Native
PG
Y
Y
9
Bidens pilosa
Exotic
AF
Y
Y
10
Polygala paniculata
Exotic
AF
Y
Y
11
Eragrostis brownii
Native
PG
Y
Y
12
Arundinella nepalensis
Native
PG
Y
Y
13
Alloteropsis semialata
Native
PG
Y
N
14
Lobelia purpurascens
Native
PF
Y
Y
15
Sonchus oleraceus
Exotic
AF
Y
Y
16
Paspalum notatum
Exotic
PG
Y
Y
17
Eremochloa bimaculata
Native
PG
Y
Y
18
Wahlenbergia gracilis
Native
AF
Y
N
19
Epaltes australis
Native
PF
Y
N
20
Schenkia australis
Native
AF
Y
N
21
Melinis repens
Exotic
PG
Y
Y
22
Velleia spathulata
Native
PF
Y
Y
23
Polygala spp
Native
Y
N
24
Phyllanthus virgatus
Native
AF
Y
Y
25
Cheilanthes sieberi
Native
PF
Y
Y
26
Paspalum urvillei
Exotic
PG
Y
N *2
27
Apiaceae spp
Exotic
PF
Y*3
Y
28
Cymbopogon refractus
Native
PG
Y
Y
29
Phyllanthus spp big
Y
N
30
Lantana camara
Exotic
S
Y
Y
31
Dianella caerulea
Native
PG
Y
Y
32
Parsonsia straminea
Native
C
Y
Y
33
Passiflora suberosa
Exotic
C
Y
N
34
Senna pendula
Exotic
S
Y
Y
35
Lomandra multiflora
Native
PG
Y
N
36
Drosera spathulata
Native
AF
Y
Y
37
Aristida queenslandicum
Native
PG
Y
N
38
Leucopogon juniperinus
Native
S
Y
N
39
Gomphrena celesoides
Exotic
PF
Y
N
40
Setaria sphacelata
Exotic
PG
N
Y
41
Sauropus hirtellus
Native
PF
N
Y
42
Scleria sp.
Native
PG
N
Y
43
Schoenus sp.
Native
PG
N
Y
44
Cyanthillium cinereum
Native
PF
N
Y
45
Cyperus sp.
PG
N
Y
46
Desmodium sp.
Native
PF
N
Y
47
Hypericum gramineum
N
Y*4
48
Panicum repens
Exotic
PG
N/Y
Y*5
Notes
*1 Awaiting seed head to confirm and id to species. Maybe D. violascens
*2 Was originally Big weed paspalum (2023)
*3 Maybe Ranunculus inundates
*4 Needs further identification
*5 Needs further identification; identified in 2023 as Cynodon dactylon in wetland area
Following on from the quail sighting (see –The Quail Turn ) the artwork sites were reconfigured as follows: The previously named ‘passive area’ or ‘passive regeneration area’ would remain as an area largely left to its own devices. Within that area, FAI site artworks would be placed that would in some way benefit localities – but it would, apart from that, remain as is – i.e. an active grassland with a healthy emerging forest cover.
The artwork ‘passive regeneration area’, June 11th (winter), 2024 (Image Keith Armstrong)
And the so called ‘active area’ – the long ‘wet gulley’ would, rather than being planted out, also be also left to self-manage – with the hope that the rare quail would return – and that grassland species would continue to favour this area – something that does seem to be happening at least with currently resident common Brown Quail. (Coturnix ypsilophora). Of course within that area we might also conduct some small growing experiments – but this approach would also conserve the soil sampling approach that had begun (See Setup (7) Further Soil-biology Adventures – from the Artwork Burn Site).
Artwork site – prior nomenclature (Image David Tucker)Artwork Site – new nomenclature – passive regeneration and wetland (Image Keith Armstrong)The artwork site situated within SERF’s overall infrastructure (Image David Tucker/SERF)
Again, this new setup was reflected in the artwork ‘Analog Intelligence’ shown at ISEA 2024 – notably with the sound of the red breasted Quail echoing through the soundtrack.
Dr. David Tucker’ standing in the artwork’s ‘wetland’ area, June 2024 (Image Keith Armstrong)
After our initial forays into the micro-biology of the soils at the artwork site, Dr. Eleanor Velasquez and I returned to A/Prof Carrie Fisher’s lab to delve further into the investigation of the health of pupureocillium bacteria. This lab investigation marked the culminating stages of A /Prof Hauxwell-led study into the effects of the burning we had done in 2023 – and how it had affected soil bacteria under the burnt areas (the area we were now calling the wet gulley or wetland). This marked the end stages of the 2nd stage of what will now be a long-term study into the effects of burning on particular soil health indicators.
Original burn in 2023 at the SERF artwork site (Image Keith Armstrong)
This therefore was time to really begin to understand what bacteria morphotypes had been identified and isolated from the soil samples across the site’s 13 transects (a feature which I also added to the loosely representational animation I was creating for ISEA 2024, as a series of stylised, subsoil ‘pots’ – see screenshot below).
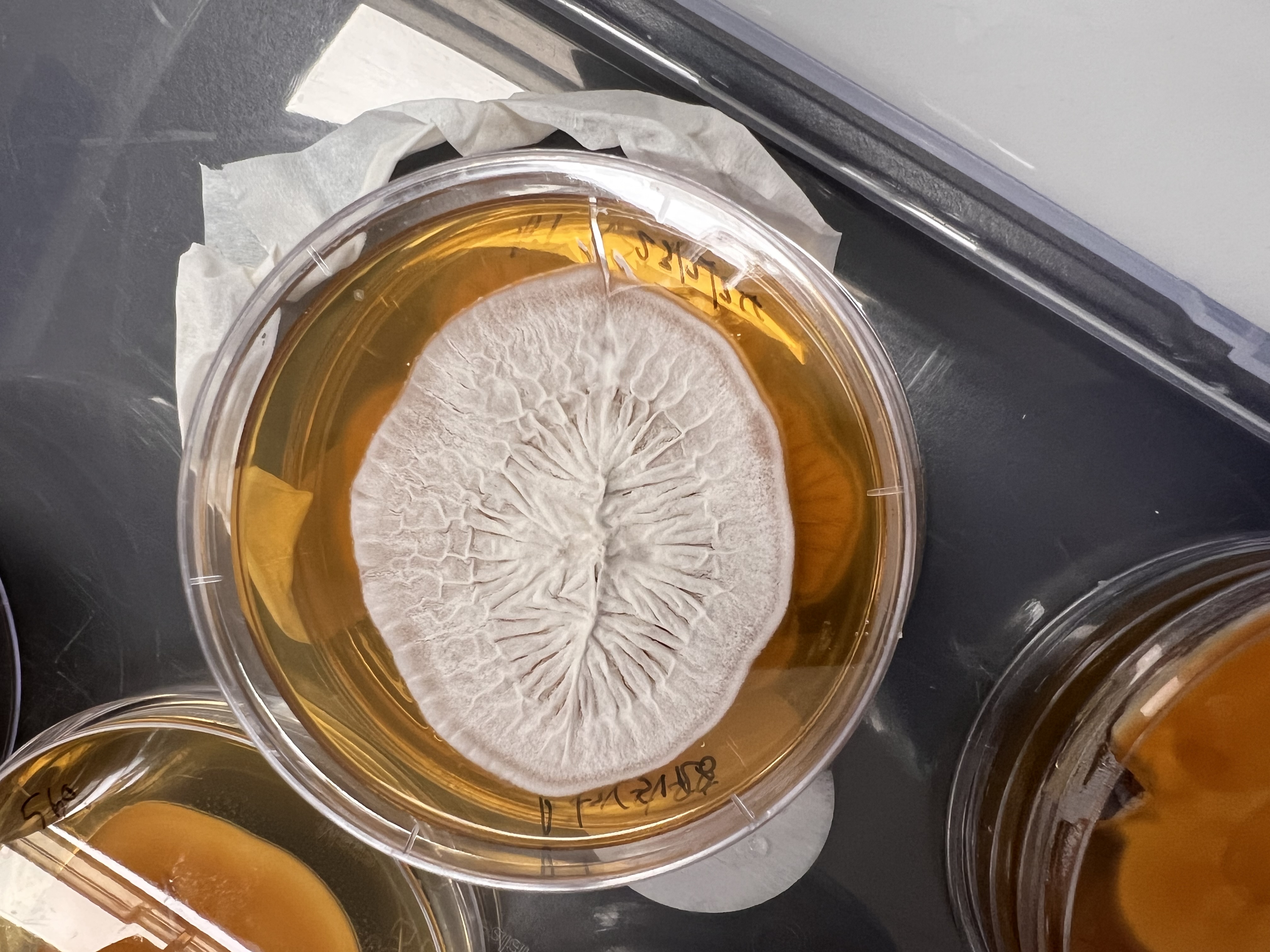
Still from the ISEA 2024 presented artwork Analog Intelligence – indicating stylised soil samples across transects (Image Keith Armstrong)Fungal Morphotype extracted and grown from the artwork site at SERF -with a wonderfully filamentous form. (Image Keith Armstrong)
Whilst this visit therefore allowed us to view the outcomes in the flesh, a couple of weeks later we were also able to listen to students present their analysis of the fungal strands that they had being attempting to identify, culture, re-test and analyse from the site – as shown in the following images:
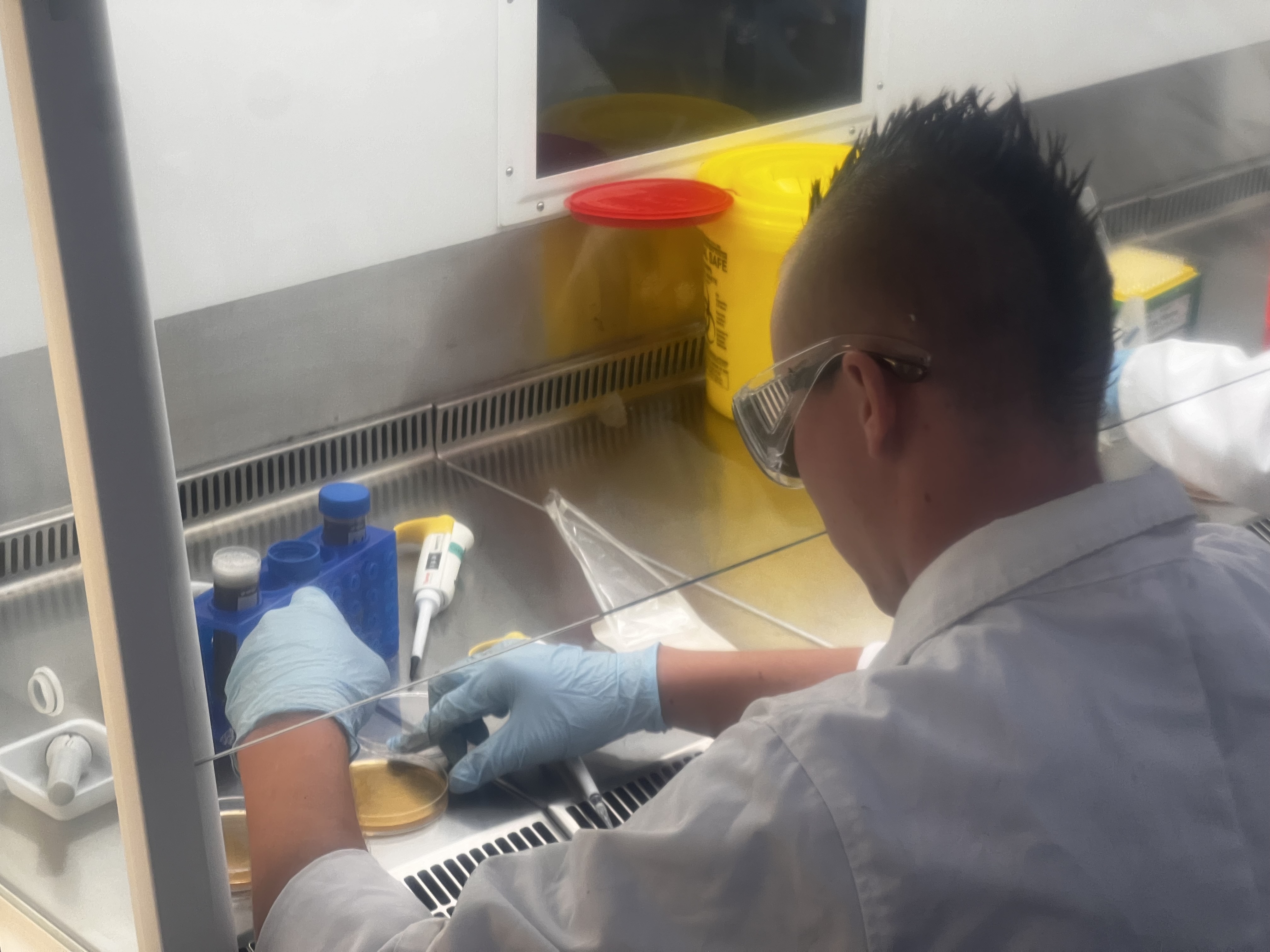
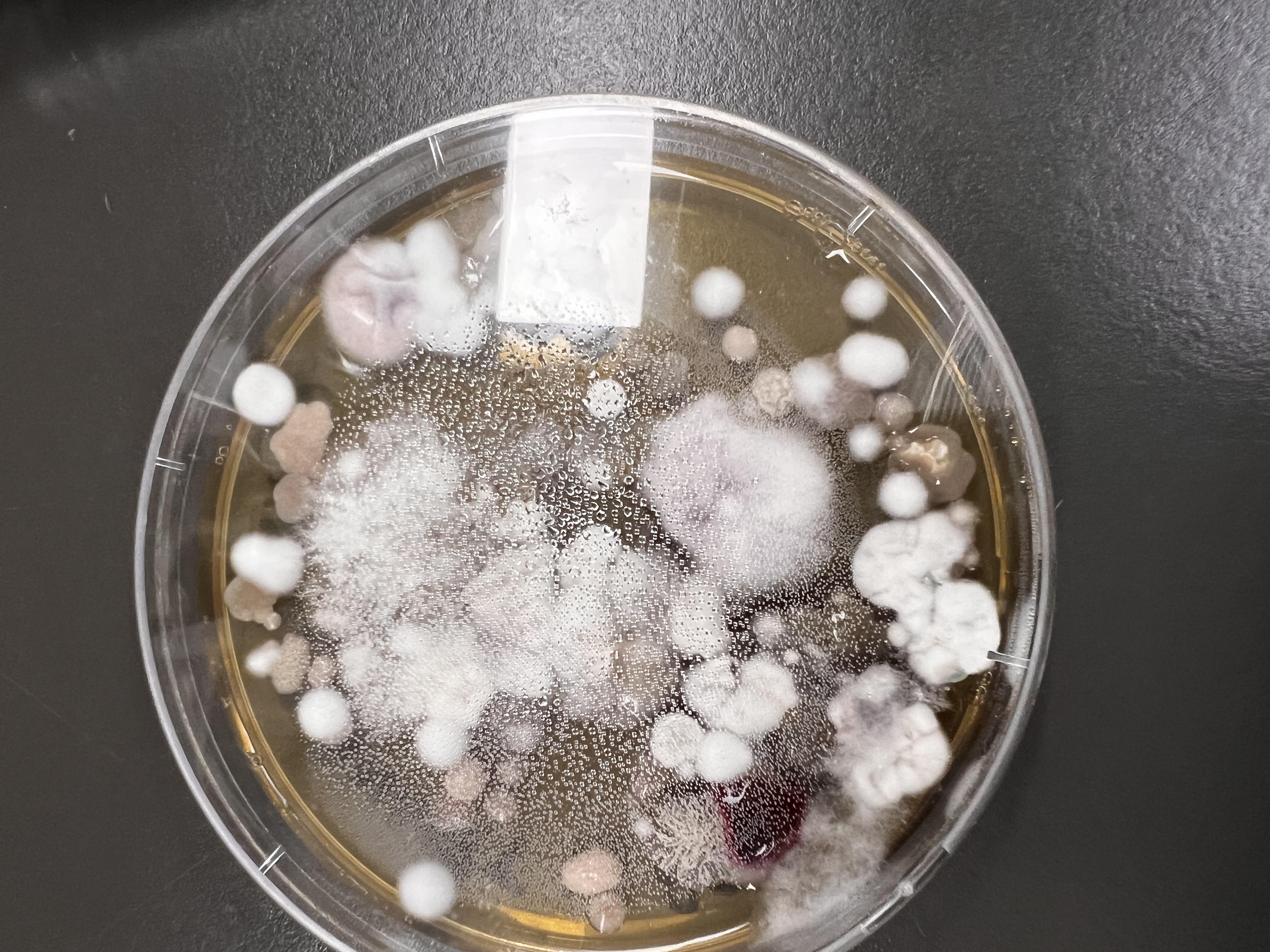
pre-prepared samples of cultured morphotypes come out of the chilling cabinets and are handed out for lab use (Image Keith Armstrong)Doing the hard yards in the lab to identify, and investigate soil bacteria (Image Keith Armstrong)Attempted fungal isolates prepared and analysed by the students from the artwork site. (Image Keith Armstrong)Details of fungal isolates from the artwork site prepared and analysed by the students. (Image Keith Armstrong)Fungal isolates and Tween – a frequently used supplement for cultivating bacteria. (Image Keith Armstrong)Comparing the plates against known/perfectly grown morphotypes supplied as a reference sheet (Image Keith Armstrong)Another filamentous bacterial form. (Image Keith Armstrong)An alternate bacterial morphotype (Image Keith Armstrong)
Presenting the Results of the Fungal Analysis
Flashback to the burn that started this whole process – the artwork site, 2023 (Image Keith Armstrong)
Results of the effects of the burn on the artwork during were subsequently presented at a student poster session to Faculty staff (with Dr. Eleanor Velasquez and myself present as guests). Dense information was presented via digital posters on large screen TVs in order to communicate outcomes – mostly in a language suited to fellow scientists. These presentations predominantly focussed on the differences between samples at the transects – i.e. whether the fungal activity differed significantly deep inside, towards the edge or on the outer of the transects (running across the artwork burnt gulley site).
Details of (anonymous) student presentation showing the background, methods, aims and type of data they had gleaned from their process (Image Keith Armstrong)Details of (anonymous) student presentation showing the background, methods, aims and type of data they had gleaned from their process (Image Keith Armstrong)
Given the number of variables – and the time the survey had been done after the burn (several months) – some student results were arguably less clear cut than perhaps we might have expected – but most showed some indications that the fungal diversity had suffered initially towards the centre of the burn site – with the assumption that it would return strongly into the future.
A so called ‘heatmap’ that indicates the abundance of bacterial morphotypes across the sample sites (Image Keith Armstrong)Still from the ISEA 2024 presented artwork Analog Intelligence – indicating stylised soil samples across transects (Image Keith Armstrong)
A Further Morphotype Workshop
Around that time Eleanor and I were also invited to participate in another ‘microbiology 101’ workshop – which we attended with interest given its hands on possibility and our need to increase our understanding of the processes involved in culturing and analyse. At this lab (led by A/Prof Hauxwell and initiated by Prof Jenn Firn (another prior collaborator from my prior Carbon_Dating project).
During this session we counted professionally pre-prepared morphotypes (a morphotype refers to the ability of certain fungi to switch between different growth forms) from agar plates with isolates (a specific strain or individual fungal soil organism that has been separated or cultured from its natural environment). This is the sort of class that’s run by the science faculty to first years a taste of biology and ecology.
The session allowed us to investigate a full set of clean, cultured reference specimens from 2 years of sampling at another local site, plus some fresh soil samples plated out for us. We then did some basic quantitative assessment of morphotypes using a guide provided – which would allow then heat mapping (two-dimensional tables of numbers as shades of colours – a popular plotting technique in biology, used to depict multivariate data) and the derivation of a Simpsons diversity index (a measure of diversity which takes into account the number of species present, as well as the relative abundance of each species).
Project collaborator Dr. Eleanor Velasquez council fungal morphotypes (Image Keith Armstrong)A type 15 Morphotype – used as a reference (Image Keith Armstrong)Expertly prepared, ‘clean’ fungal morphotypes as references, prepared by A/Prof Hauxwell’s team) (Image Keith Armstrong)Expertly prepared, ‘clean’ fungal morphotypes as references, prepared by A/Prof Hauxwell’s team) (Image Keith Armstrong)Project scientist Dr. Eleanor Velasquez council fungal morphotypes (Image Keith Armstrong)Expertly prepared, ‘clean’ Type C fungal morphotype reference, prepared by A/Prof Hauxwell’s team) (Image Keith Armstrong)
Developing a Timelapse of Fungal Growth
One of A/Prof Masters students Edward Bryans, assisted by Genevieve Dickson very kindly agreed to setup a time-lapse for us so we could see the development of these bacteria across time.
QUT Researcher Edward Bryans examining (exotic) Buffel grass infected with mealy bugs (Image Keith Armstrong)
Time lapses are a bit of a staple of science shows, but trying to make your own is a totally different thing as it becomes an ’embodied’ experience. If you watch someone’s else’s – you don’t really know how long it took – but making it yourself, you have the experience of the actual bacteria that you’ve sighted – that can ultimately become compressed into those 30 seconds. You share life with that thing that you’ve been photographing, and by embodying it, it becomes a totally different way of experiencing and understanding the world that more than human is a part of.
Edward and his colleague Genevieve built a simple system in an airflow protected cabinet in their lab using a 35mm camera with protective coverings – so that we could monitor the bacteria in action. The process turned out to be quite a fiddle – with the heat of the lights drying out the fungal plates and restricting growth – so it’s an ongoing project!
QUT Researcher Edward Bryans showing a preserved fungal growth plate (Image Keith Armstrong)QUT Researcher Edward Bryans at the cold cabinet preserving fungal growth plates (Image Keith Armstrong)The time lapse setup for recording final growth of purpureocillium in a hot cabinet. (Image Keith Armstrong)
This is the first outcome – showing the purpureocillium and a rather more aesthetic contamination yeast moving in unexpectedly! More to come as Edward and Genevieve work out an improved method to get a larger more impressive growth 🙂
Intelligence is not something to test – but rather to recognise ..
The Quail Turn: A Project Pivot
EXCITINGLY in late April a rare Red backed button quail (Turnix maculosus) had been sighted by the SERF bird group/Peter Storer. These birds are rarely seen and poorly studied – being agents of disguise within their preferred wet grassland environments – with a call that is a soft repeated ‘oom-oom-oom-oom’ and with the end of each note ascending in pitch. They are most often seen breaking wildly for cover when disturbed and flying elsewhere in the grassland. They are listed as vulnerable (to extinction in NSW), although not in Qld. The itinerant ones are thought to arrive Oct/Nov and likely leave for the North late Feb/March – but this year’s wet hot season has likely kept them longer. (It is likely that the birds had in this case left end of April – pre the slashing season at SERF which is later than normal).
Rare Red-backed button quail (Turnix maculosus) sigthed in the artwork site (Image Peter Storer)
By chance I met Peter Storer in the paddock with his binoculars looking for the quail in late April using call sounds – as I was working late in the afternoon on setting up key sites. During that time at dusk, when they are noted to be active. At that time I spotted a dead juvenile red backed button quail at the edge of the long grass – indicating breeding had happened (they build nests in shallow depressions in the grasses) .. suggesting that great care should be taken entering any of the artwork’s long grassed areas esp. during nesting season.
Deceased Red-backed button quail at the artwork site (Image Keith Armstrong)
Given that the site has been in essence maintained as a grassland by slashing for decades – and that this had attracted a rare find – this raised questions as to whether regenerating the previously burnt gulley/regeneration area with trees, or indeed premature slashing of other parts of the property, may drive them away/destroy their nests/cause them to not stop in the area in the future?
Clearly this finding presaged a project turning point of some kind that would require consultation across a number of groups and the science team. It seemed that the idea of ‘active’ and ‘passive’ areas may now need to change. And that eerie, profound, quail ‘oom-oom-oom-oom’ – it may well resonate across the future artwork ..
FYI .. other birds know to be at the site (ref. Peter Storer) were the Brown Quail (Coturnix ypsilophora) which is quite common and likely resident, the Golden-headed Cisticola (Cisticola exilis), Tawny Grassbird (Cincloramphus timoriensis), and Sacred Kingfisher (Todiramphus sanctus).
During April/May 2024, consistent with SERF’s other monitoring programs, and the need to track progress scientifically on this project, I decided to initially establish 1-2 trail cameras on site – as phenocams (vegetation change cameras) – and acoustic monitoring.
Acoustic Monitoring I determined in consultation with Dr. David Tucker that initially 1 Solar powered BioAcoustic Recorder (Solar BAR) should be deployed near the bottom of the passive plot where it meets the active plot – to record continuous audio data for later analysis – given these devices’ detection radii is quite wide on open pasture (>100m):
Image of the site showing fungal transects, the position of the acoustic recorder and proposed future artwork sites (Image Keith Armstrong)
The choice of this hardware is consistent with the equipment used by the Australian Acoustic Observatory which Dr. David Tucker is a member of – ensuring he will be able to help identify the call sounds.
Acoustic Spectrogram of Bird Sound (Image courtesy of Frontier Labs)
SolarBAR recorder + Dr. David Tucker and Prof Paul Roe (Australian Acoustic Observatory)
The initial acoustic monitor was installed on June 11th 2024.
Dr. David Tucker installing the acoustic monitor settings, June 11, 2024 (Image Keith Armstrong)
Visual Monitoring
Phenocams are digital cameras (usually trail cams more typically used to capture nocturnal animal movements) set up to capture photo time-lapse images of foliage (ranging from one per 30 mins to one per day) at the same times. Scientists use these continuous visual records to observe things like vegetation development, including flowering, fruiting, and leaf lifecycles. I this way they are able to generate quantitative measures of plant phenology (timings of cyclical or seasonal biological events, that might include flowering, migrations, egg laying or hibernation).
4.0cg Sy electronic Technical co ltd Trail Cam with 4G – one of the various cameras I have to hand
Aerial Monitoring This would be a mixture of input from the QUT REF team who fly 35mm quality cameras – and my own Mavic 2 Zooms. Initial photos were taken on 24/4/24 showing some of the area put aside for the artwork.
A part of the artwork site, April 2024 (Image Keith Armstrong)
Dr. Hauxwell has been using adjacent areas at SERF and part of the artwork/revegetation site for her research experiments into beneficial soil fungi – which have an end outcome to reduce pasture dieback that is caused by mealy bugs. (The bug in question is the paspalum mealybug, Heliococcus summervillei – and her work is of particular relevance to the pasture industry given that these invasive bugs devour Buffalo grass which is a key, commercial pasture grass, (and interestingly a notable environmental weed in conservation contexts). To do this work Caroline and her research team have become experts in isolating fungi from soils of forests and pastures.
Collecting soil samples along transects at SERF – across the artwork site (Image Keith Armstrong)
Immediately before, and following the burn of the SERF wet gulley/artwork site A/Prof Hauxwell had initiated a periodic soil sampling regime to learn how the fungal composition of the site might change according to the burn – something which also seemed very relevant to us given we needed proxy ways to determine the improvement of soil and plant health as the artwork process evolves.
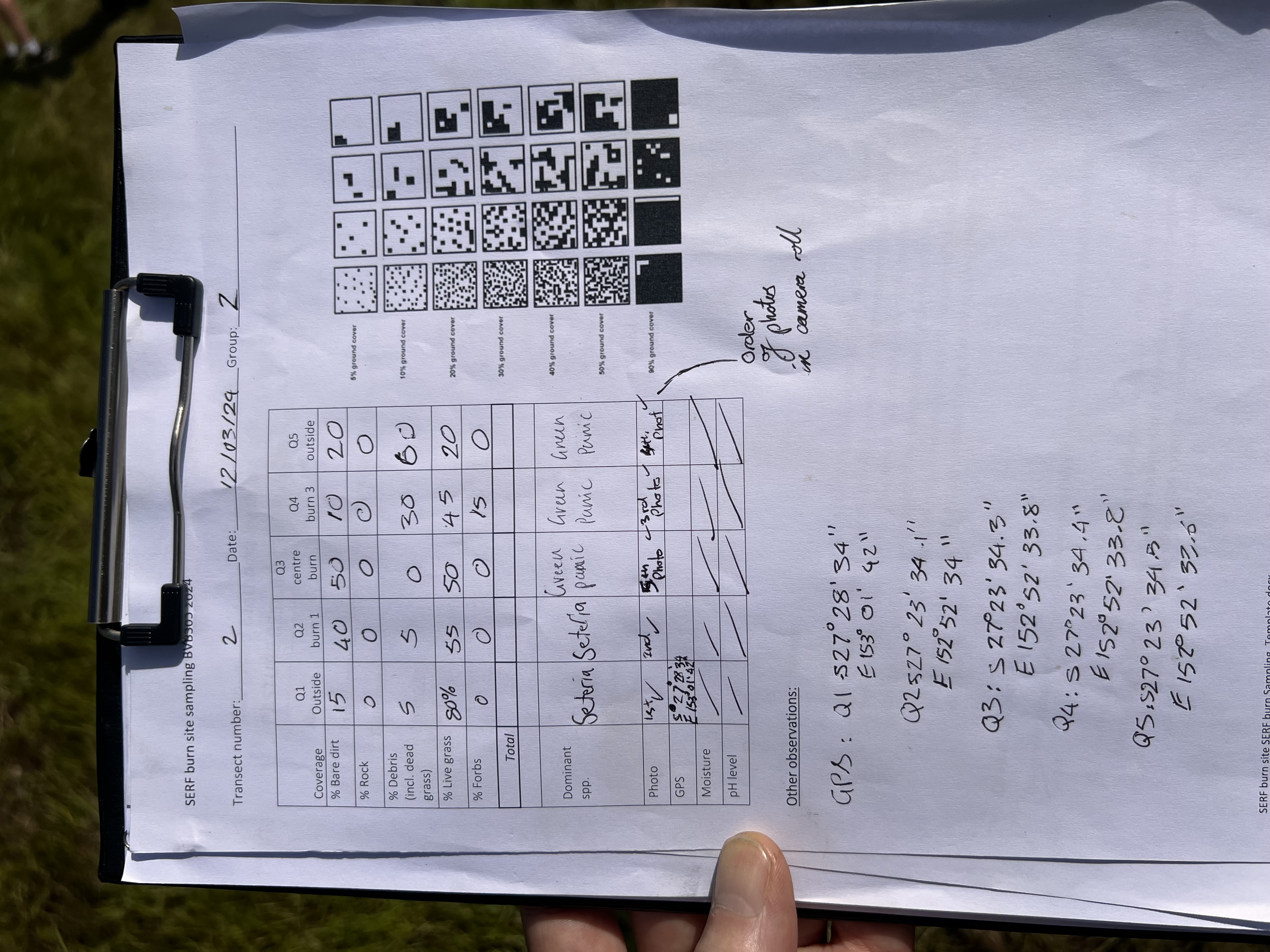
So on 11/3/24 Dr. Eleanor Velasquez and I joined the soil sampling process at SERF – to understand and observe more about the scientific methods underway.
Soil samples from the artwork zone, collected and kept cool (Image Keith Armstrong)Soil samples from the artwork zone (Image Keith Armstrong)Artwork/site survey team, March 2024 (Image Keith Armstrong)Site sampling notation (Image Keith Armstrong)
As per the prior study, samples would be taken in transects across the whole site (5 points per transect spanning the strip that we burned last year). Her students and postgraduate team had been analysing those soil samples for the presence of Purpureocillium (see prior soil biology investigation post).
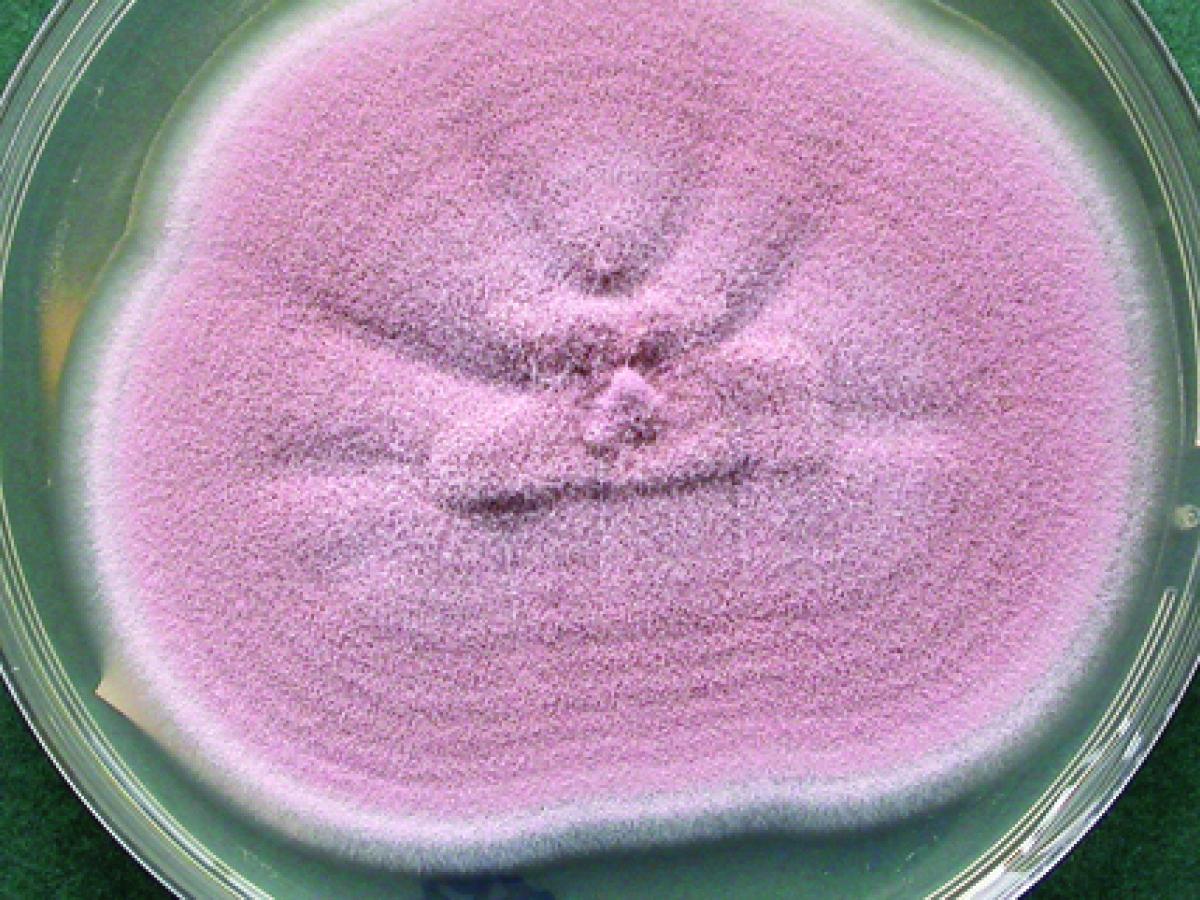
Purpureocillium lilacinum is commonly isolated from soil, decaying vegetation, insects, nematodes and as a laboratory contaminant. It is also a causative agent of infection in human and other vertebrates (Luangsa-ard et al. 2011).
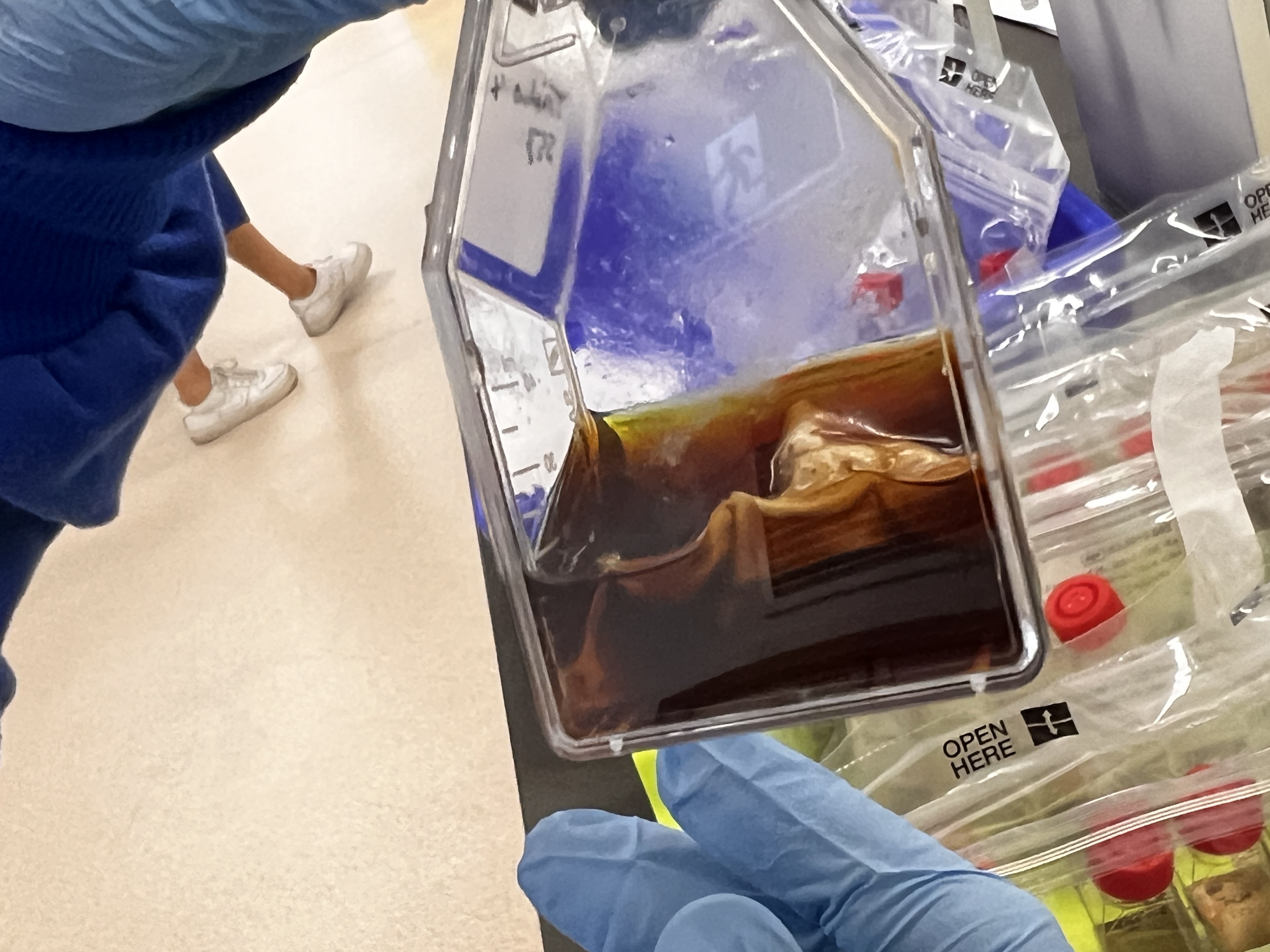
Purpureocillium lilacinum (Subcultures from broth) (Image Keith Armstrong)
During our discussions we learnt that she would be continuing this work into 20924 – and she invited Eleanor and I to join her class which we have now done on a few occasions. The intentions of her study were to pursue three themes ..
Soil Ecology: The isolation of facultative root endophytes, particularly P. lilacinum, to determine the effects of burning on abundance and diversity, and diversity within P. lilacinum.
Media Development: Development of media for the production of P. lilacinum variants as an inoculum (that is – a liquid solution of the fungi that roots can grow through to make them resistant to Mealy bug) – which leads to the third area of interest
The Good Bugs: The application of P. lilacinum variants as an inoculant against pasture mealybug.
Keith And Dr. Eleanor Velaqsquez in the soil biology lab, March 2024 (Image Keith Armstrong)
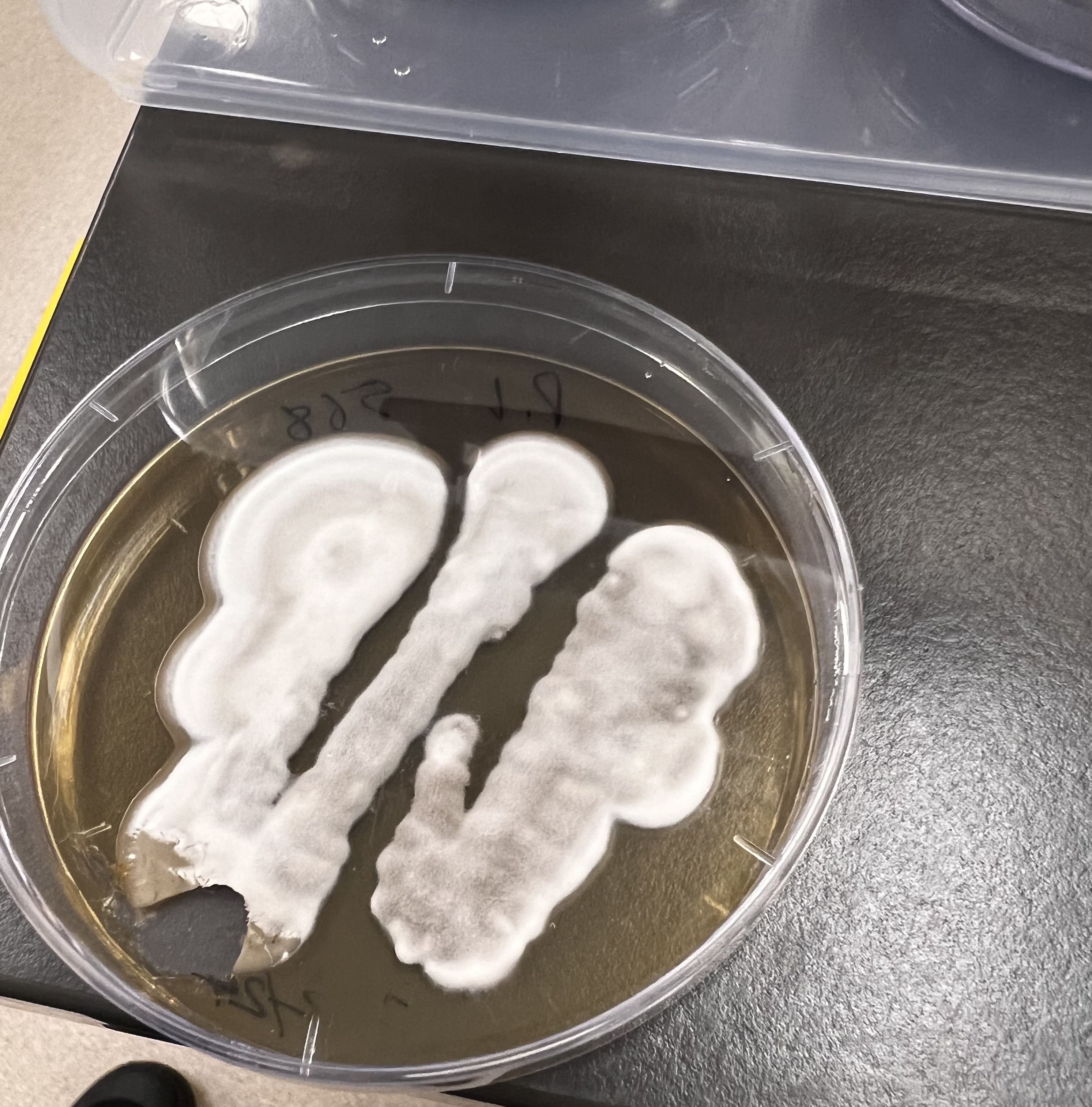
For me it has been a fascinating return to the analogue chemistry methods I remember only from Year 10 (!) – and has included both sampling soil to the required protocol at SERF and then observing its analysis, culturing and the isolation of so called ‘morphotypes’ (in essence any of a group of different types of individuals of the same species in a population/individual fungi) for further growth on agar plate cultures. (An agar plate is a petri dish that contains a growth medium solidified with agar, and is used to culture microorganisms).
Student preparing soil solution containing bacteria onto an Agar plate (Image Keith Armstrong)Soil fungi experiments with A/Prof Caroline Hauxwell, 2024 (Image Keith Armstrong)
Inoculation medium/broth with dissolved fungi – ready for applying to grass plant roots (Image Keith Armstrong)A mixture of different fungi to be described and isolated. (Image Keith Armstrong)Notes on the composition (numbers of morphotypes and characteristics) of the fungi cultured over the past week – recorded by a student (Image Keith Armstrong)
The descriptions alone are evocative and the structure, form and ‘intelligence’ of this form suggest fruitful investigation ahead!
Preparing agar plates for the second stage of growth (Image Keith Armstrong)
The exciting idea that Caroline has raised – which further cements the fruitful connections emerging between the arts and sciences, is “to do a longitudinal study of the soil microbiome by adapting the sampling this year (across the burn site) to compare the dryer slope with the burned gully below.
Her (and our) interest would therefore be to see the changes in soil fungal diversity over time, particularly once trees are established.” At this stage she is looking to do this highly technical, lab based work over several years – which will allow this project an extraordinary look into the health of the soil as the artwork develops. Exciting times ahead – so many thanks to A/Prof Hauxwell for your kindness, interest and engagement on this co-beneficial process 🙂
Purpureocillium lilacinum (Subcultures from broth) (Image Keith Armstrong)
Following discussions with the team – notably Gabrielle Lebbink and Eleanor Velasquez – we began to open up a new thread around the possibilities of soil inoculation – that is – finding ways to move material from eco active parts of the existing forest to the new sites. This investigation is fuelled by/consistent with the need for the design of the artwork work to be of positive net benefit to the ecology of the emerging forest.
Gabrielle supplied some great information about ‘Re-wilding microbial/soil based communities’:
Contos P, Wood JL, Murphy NP, Gibb H. Rewilding with invertebrates and microbes to restore ecosystems: Present trends and future directions. Ecol Evol. 2021;11:7187–7200. https://doi.org/10.1002/ece3.7597. Downloaded y on [02/02/2024].
This article offered us some pointers of how we may be able to enhance soil biodiversity (potentially a part of an art/science process of trslocation of materials from the current forest to teh regeneration zone). I noted of that paper ..
Re-wilding of Microbe Communities (Notes, KMA 7/2/24)
Notes from: Contos P, Wood JL, Murphy NP, Gibb H. Rewilding with invertebrates and microbes to restore ecosystems: Present trends and future directions. Ecol Evol. 2021;11:7187–7200. https://doi.org/10.1002/ece3.7597. Downloaded from https://onlinelibrary.wiley.com/doi/10.1002/ece3.7597 by National Health and Medical Research Council, Wiley Online Library on [02/02/2024].
CONTEXT: Invertebrates/microbes are key drivers of landscape scale ecosystem functions such as nutrient recycling and carbon sequestration, but many of these may still be functionally absent decades after a restoration process. The aim of any restoration project should actually be to reinstate ecosystem function and reestablishment of self-organising communities; and yet restoration ecologists rarely consider invertebrates and microbes which these make up the vast bulk of biodiversity and may drive key ecosystem recovery processes. Currently there is limited use of this approach in re-wilding projects – and often poor understanding of its importance.
Stepwise restoration – i.e. a species at a time – is becoming less realistic in a dynamic and rapidly changing world. Typical approaches to restoration involve plant only and plant and animal only approaches – which operate under what the authors call the ‘field of dreams paradigm’ – i.e. that if you build the habitat they will come! Such attempts often therefore ignore the ‘unseen majority’.
Normally microbial species translocations are done for the benefit of other species (i.e. to encourage back butterflies or for the benefit of a particular plant health)) rather than for themselves per se (e.g. introduced AMF fungi) – without considering whole communities. Furthermore, in most cases microbes are rarely monitored afterwards anyway. Hence some environments may remain functionally unrecovered 10 or 20 years later. NB The process of microbial re-wilding is more common in soil inoculation studies (soil transplants).
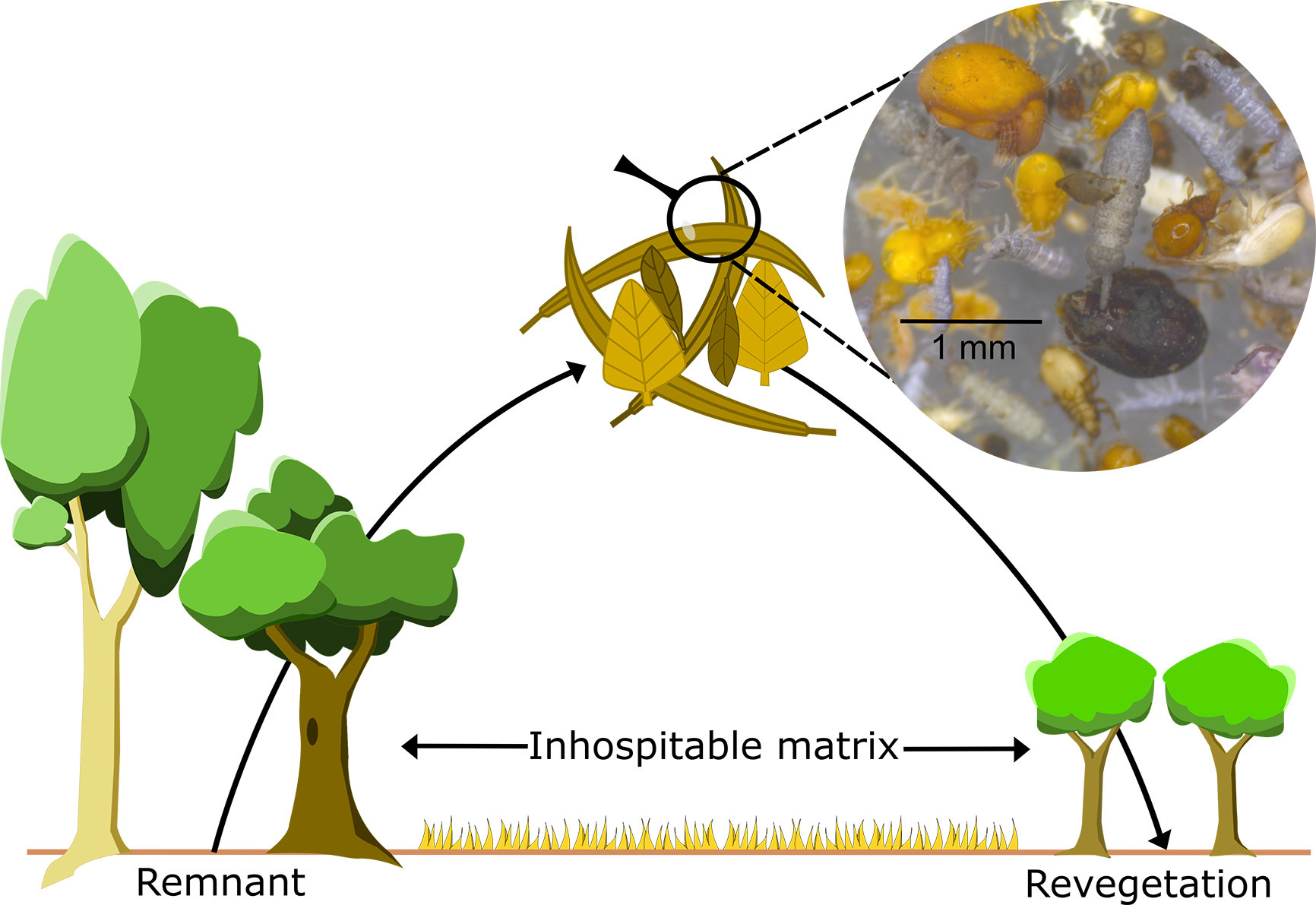
Diagram from paper showing how we might move material across the ‘Inhospitable Matrix’ that bounds the artwork site: “Leaf litter samples taken from remnant patches and moved into revegetation patches will carry a multitude of invertebrate and microbe species and individuals. Inset: detritivorous mites and springtails taken from a leaf litter sample”.
Whole of community microbial (and invertebrate?) rewilding is often practical due to ease of manipulation – e.g. in forest litter invertebrates and microbes (e.g. detritovores) are critical components – (see p7193). Whole of community rewilding typically involves transporting small subsets of whole habitats – normally from a nearby remnant site to a revegetation plot that maybe separated by an ‘inhospitable matrix’ – e.g. a pasture – and may include elements such as soil, dead wood and pond mud.
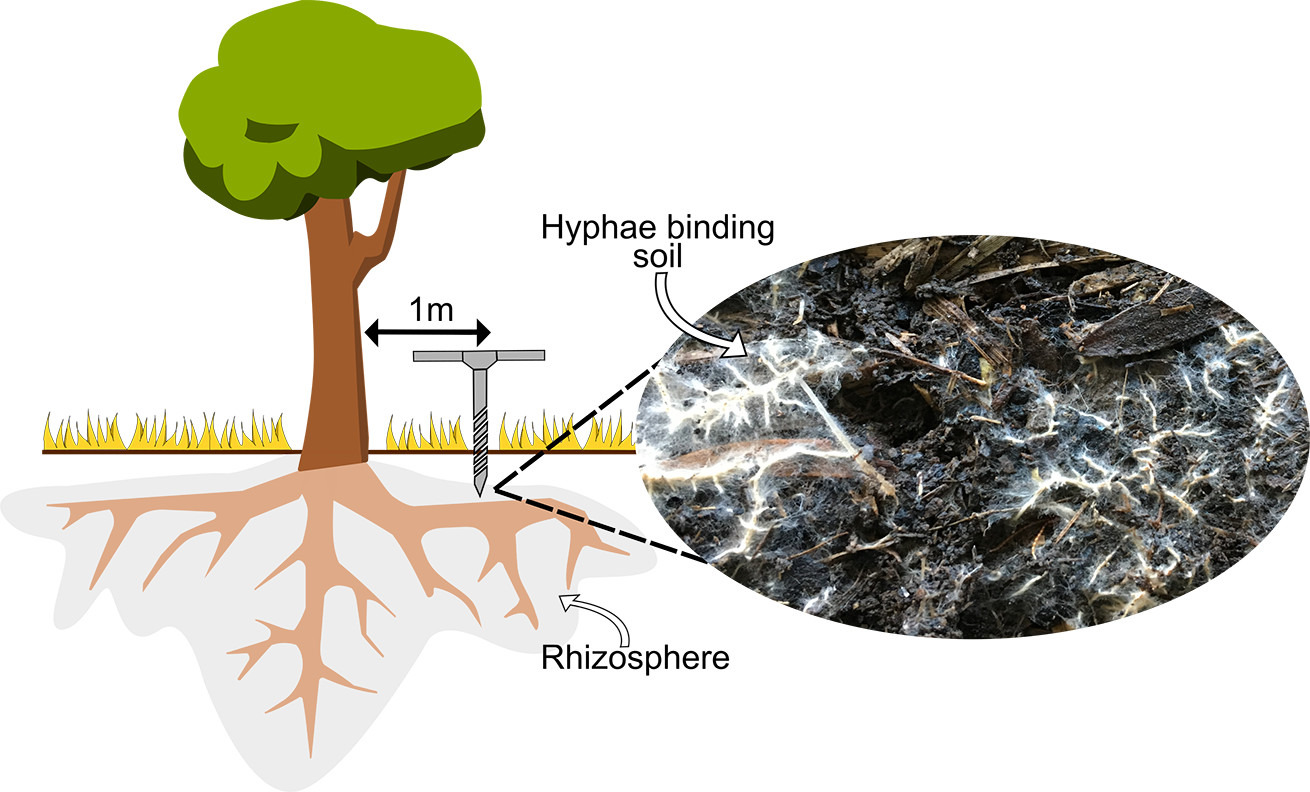
“Collecting soil from the whole rhizosphere region of large established trees is impractical. Collecting rhizosphere communities by sampling 1 m out from the base of trees using a soil corer is a viable methodologic approach and is a more targeted way of rewilding microbial communities than current soil inoculation studies. Previous research has demonstrated that rhizosphere signatures can be detected using this approach for microbe communities from rainforest plant species despite the complex overlapping root networks (Wood et al., 2020)”
Its critical to avoid failures which will result in overall net loss of biota in an already stressed environment). The authors therefore suggest considering:
suitability of renewed habitat to accommodate the microbes: The revegetation site should be ameliorated and receptive (physical and chemical limitations of soil) – and the source site very carefully scanned for invasives and pathogens.
dispersal limitations of some invertebrates and microbes
Distance of new site from existing vegetation ideally should not be far
May work better in cool wet conditions when microbes and invertebrates are active and closer to surface of litter level (n.b. authors outside Australia)
Broadly speaking a project should
Set restoration goals
Evaluate the restoration trajectory
Choose what to re-wild
Conduct post re-wilding monitoring
Evaluate success/failures.
“Litter communities contain a breadth of species, including trilobite cockroaches, Laxta granicollis (center), and armadillid isopods (top right). These taxa are often overlooked during rewilding projects, despite their immense contribution to biodiversity and their influence on ecosystem functions such as decomposition. Photo credit: L Menz”
These interesting learnings dovetailed with the research one of the other science academics (A/Prof Caroline Hauxwell) who is investigating diversity of fungal communities in soil, including Purpureocillium, which is a interesting fungus associated with pasture health and resilience to pests and diseases. She is interested in the impacts of burning on beneficial fungal symbionts (and thus the artwork site is an ideal choice) – something that ties in with her work on management of the pasture mealybug Heliococcus summervillei, which causes ‘pasture dieback’ (research that is co-funded with Meat and Livestock Australia). A/Prof Hauxwell is one of Australia’s foremost experts in this area and her research is extensive and widely recognised.
Dr. Hauxwell had already independently set up an experiment to monitor the artwork areas pre and post burn – so we were all ears to her process! Generously she invited Eleanor and I to join her class and get involved in some of her soil science – we jumped at this opportunity as an interesting creative opening.
To get more of a handle on this work – here are some excerpts from an email that A/Prof Caroline Hauxwell subsequently wrote – as part of a group email in Feb 2024.
We welcome, Keith Armstrong, who initiated the burn at SERF and who is very interested in this work. Welcome, Keith.
“For most of you the work really starts in week 3 with the field trip to collect soil samples at SERF.
The project work this year is a Deep Dive on Soil Endophytes. We’ll be focusing particularly on Purpureocillium lilacinum. The project builds on sampling that we did before and shortly after a burn site at SERF for capstone last year.
The student data identified changes in Purpureocillium abundance and distribution in the burned area.
Fungal culture (Image c/o Sean Martin, created with particiapnts at the 2021 Woodford Bio Lab)
We also know from our research that P. lilacinum isolates have significant morphological diversity, but ITS sequencing has shown only 1 sequence/variant. We hope to look a bit more closely at P. lilacinum diversity in this project. All groups will also have access to 3 PCR primers for Sanger sequencing: one to characterise fungi to genus (ITS) and 2 others to take a deep dive on the identity of P. lilacinum morphotypes. For reference see Naimul’s work on Metarhizium. (https://eprints.qut.edu.au/view/person/Islam,_Shah.html) and https://doi.org/10.1016/j.funeco.2022.101179.
Soil Ecologists: The isolation of facultative root endophytes, particularly P. lilacinum, the effects of burning on abundance and diversity, and diversity within P. lilacinum.
Media Types: Development of media for the production of P. lilacinum variants as an inoculum
Fungal culture (Image c/o Sean Martin, created with participants at the 2021 Woodford Bio Lab)
All students (plus artists and the science team) will take part in soil sampling .. at SERF. This will take soil samples at 5 points along a transect across a strip that was burned last year See picture attached). 2 sample points will be outside the burn, and 3 inside it. All will plate out their soil samples onto selective media.
For further background of this process see a report of the 2021 lab run by A/Prof Caroline Hauxwell at Woodford





















































































{kind=link}